
User:Vtcell/sandbox
Paracytophagy (from Ancient Greek para 'nearby' kytos 'cell' and phagy 'eating') is the cellular process whereby a cell engulfs a protrusion which extends from a neighboring cell and may contain material which is actively transferred between the cells.
Introduction
[edit]The process of paracytophagy was first described as a crucial step in the life cycle of the bacterial pathogen Listeria monocytogenes. Paracytophagy allows Listeria and other intracellular pathogens to spread directly from cell to cell, thus escaping immune detection and destruction. Studies of this process have contributed significantly to our understanding of the role of the actin cytoskeleton in eukaryotic cells.
Actin cytoskeleton
[edit]Actin nucleation factors
[edit]Although previously the cell movements in pseudopods were though to be a simple conversion from gel to sol and vice versa, nowadays it is well understood that cells emitting pseudopods or lamellipodia depends on actin for these processes. Actin is one of the main cytoskeletal proteins in eukaryotic cells. The polymerization of actin filaments are responsible for cell movements like pseudopods, filopodia and lamellipodia.
Actin polymerization at the leading edge
[edit]Cells actively builds actin microfilaments that push the cellular membrane towards the direction of advance. The actin monomers are preferentially assembled on the leading (distal) edge and disassembled on the delayed (proximal) end, the same units are transported towards the leading edge to conform the growing filament in a process called treadmilling.[1]
The ARP2/3 complex and nucleation promoting factors (NPFs)
[edit]Nucleation factors are enhancers of actin polymerization and as the name says, these factors contribute to trimeric polymerization nucleus formation, a structure required to start the process of actin filament polymerization in a stable and efficient way. Nucleation factors like WASP protein acts by bringing together ARP2 and ARP3 to form the ARP2/3 nucleation complex. Actin related Proteins 2/3 (ARP2/3) ensemble in a complex of seven proteins that is able to cap the minus end of actin, allowing faster polymerization of the plus end, and filament branching. [2]
WASP analogues used by pathogens to move through the host cell cytosol
[edit]Some intracellular pathogens like Listeria and Shigella, manipulate host cell actin polymerization to move in the cytosol and spread to neighboring cells. Further insight on the function of WASP protein has been enlighten by studding these bacteria, especially Lysteria ActA protein. ActA is a nucleation promoting factor that mimics WASP and is expressed in one end of Listeria, allowing ARP2/3 nucleation in there and pushing the bacteria in the opposite direction, leaving an actin comet tail behind. Other pathogens like Shigella also moves using actin polymerization and leaving a comet tail behind. In the case of Shigela, this bacterium uses a protein that recruits host cell WASPs to one end of the bacteria, allowing the microorganism to move in the opposite direction. [1], [2]

Exchange of cellular material between adjacent cells
[edit]It had been reported that filopodia-like protrusions directed toward neighboring cells in a culture of rat PC12 cells are capable of transport organelles and vesicle coated structures by means of tunneling nanotubes [3].
Sub-cellular organelle transfer (the case of Mitochondria and antigen containing vesicles)
[edit]Among the antigen containing vesicles is particularly important the case of trogocytosis, where lipids rafts or membrane patches are exchanged between immune cells in order to respond against a foreign stimuli [4]. Moreover, multivesicular compartment containing smaller vesicles into a larger vesicles called exosomes, had been shown to deliver not only antigens for cross-presentation[5], but also MHCII and co-stimulatory molecules for lymphocyte T activation [6]. In non-immune cells it has been demonstrated that mitochondria can be exchanged intercellularly to rescue metabolically unviable cells [7], a finding that was also supported recently in cancer cells [8]
Intercellular transfer by endocytocis as a niche signaling
[edit]Close contact between different cell types is necessary for bone marrow homing. During the process, cells of the surrounding bone engulf pieces of bone marrow hematopoietic cells. These osteoblasts make contact with hematopoietic stem-progenitor cells (HPC) by means of nanotubes, and pieces of the donor cells (HPC) are transferred over time to various endocytic compartments of the osteoblasts [9].
Argosomes and melanosomes
[edit]Argosomes are derived from basolateral epithelial membranes and allow communication between adjacent cells. They were first described in Drosophila, where they acts as a vehicle for the spread of molecules through epithelia of imaginal discs [10]. Melanosomes are also transferred by filopodia from melanocyte to keratinocytes. It involves classical filopodial forming pathway, with Cdc42 and WASP as key factors [11]. Argosomes, melanosomes and other examples of epithelial transfer had been compared with Listeria paracytophagy phenomenon, which can be viewed as a special case of intercellular material transfer between epithelial cells [3].
Role in the life cycle of intracellular pathogens
[edit]The two main examples of paracytophagy are the cell-cell transmission of Listeria monocytogenes and Shigella flexneri. In the case of Listeria, the process was first described in detail using electron microscopy[12] and video microscopy[13]. The following is a description of the process of cell-cell transmission of Listeria monocytogenes, primarily based on Robbins et al. (1999)[13]:
In an already infected "donor" cell, the bacterium expresses ActA, which results in formation of the actin comet tail and movement of the bacterium throughout the cytoplasm. When the parasite encounters the donor cell membrane, it will either ricochet off it or adhere to it and begin to push outwards, distending the membrane and forming a protrusion of length 3-18 μm. The close interaction between the bacterium and the host cell membrane is thought to depend on Ezrin, a member of the ERM family of membrane-associated proteins. Ezrin attaches the actin-propelled bacterium to the plasma membrane by crosslinking the actin comet tail to the membrane, and maintains this interaction throughout the protrusion process [14].
-
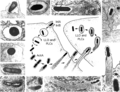 Caption1
Caption1 -
 Caption2
Caption2
_409-14,_Figure_1.png)
As the normal site of infection is gut columnar epithelium, cells are packed closely together and a cell protrusion from one cell will easily push into a neighboring "target" cell (without rupturing the target cell membrane or the donor protrusion membrane yet). At this point, the bacterium at the tip of the protrusion will begin to undergo "fitful movement" caused by continuing polymerization of actin at its rear. After 7-15 minutes, the donor cell membrane pinches off and fitful movement ceases for 15-25 minutes due to depletion of ATP. Subsequently, the target membrane pinches off (taking 30-150 seconds) and the secondary vacuole containing the bacterium forms inside the target cell cytoplasm.
Within 5 minutes, the target cell finally becomes infected when the secondary vacuole begins to acidify and the inner (donor cell-derived) membrane breaks down through the action of bacterial phospholipases (PI-PLC and PC-PLC). Shortly thereafter, the outer membrane also breaks down as a result of the action of the bacterial protein listeriolysin O[15] which punctures the vacuolar membrane. A cloud of residual donor cell-derived actin persists around the bacterium for up to 30 minutes. The bacterial metalloprotease Mpl cleaves ActA while the bacterium is still within the acidified secondary vacuole in a pH-dependent fashion, but new ActA transcription is not required as pre-existing ActA mRNA can be utilized to translate new ActA protein. The bacterium finally regains motility and the infection proceeds.
| Key Stages of Paracytophagy during Listeria infection |
|---|
| Actin motility and membrane protrusion formation using host cytoskeletal elements |
| Uptake into target cell and formation of a double membrane secondary vacuole |
| Vacuolar membrane dissolution and infection of target cell |
See also
[edit]The process of paracytophagy is considered distinct from similar but unrelated processes such as phagocytosis and trogocytosis. Some related concepts include:
References
[edit]- ^ a b Molecular Biology of the Cell. 4th edition. Alberts B, Johnson A, Lewis J, et al. New York: Garland Science; 2002.
- ^ a b Cellular Microbiology, 2nd Ed., edited by Pascale Cossart, Patrice Boquet, and Staffan Normark and Rino Rappuoli.Washington, DC: ASM Press; 2005.
- ^ a b Rustom, A.; Saffrich, R.; Markovic, I.; Walther, P.; Gerdes, H. (2004). "Nanotubular Highways for Intercellular Organelle Transport". Science. 303 (5660): 1007–1010. Bibcode:2004Sci...303.1007R. doi:10.1126/science.1093133. PMID 14963329.
- ^ Ahmed, K. A.; Munegowda, M. A.; Xie, Y.; Xiang, J. (2008). "Intercellular Trogocytosis Plays an Important Role in Modulation of Immune Responses". Cellular and Molecular Immunology. 5 (4): 261–269. doi:10.1038/cmi.2008.32. PMID 18761813.
- ^ Testa, J. S.; Apcher, G. S.; Comber, J. D.; Eisenlohr, L. C. (2010). "Exosome-Driven Antigen Transfer for MHC Class II Presentation Facilitated by the Receptor Binding Activity of Influenza Hemagglutinin". The Journal of Immunology. 185 (11): 6608–6616. doi:10.4049/jimmunol.1001768. PMID 21048109.
- ^ Théry, C.; Duban, L.; Segura, E.; Véron, P.; Lantz, O.; Amigorena, S. (2002). "Indirect activation of naïve CD4+ T cells by dendritic cell–derived exosomes". Nature Immunology. 3 (12): 1156–1162. doi:10.1038/ni854. PMID 12426563.
- ^ Spees, J. L.; Olson, S.; Whitney, M.; Prockop, D. (2006). "Mitochondrial transfer between cells can rescue aerobic respiration". Proceedings of the National Academy of Sciences. 103 (5): 1283–1288. Bibcode:2006PNAS..103.1283S. doi:10.1073/pnas.0510511103. PMC 1345715. PMID 16432190.
- ^ Rebbeck, C. A.; Leroi, A. M.; Burt, A. (2011). "Mitochondrial Capture by a Transmissible Cancer". Science. 331 (6015): 303–303. Bibcode:2011Sci...331..303R. doi:10.1126/science.1197696. PMID 21252340.
- ^ Gillette, J. M.; Larochelle, A.; Dunbar, C. E.; Lippincott-Schwartz, J. (2009). "Intercellular transfer to signalling endosomes regulates an ex vivo bone marrow niche". Nature Cell Biology. 11 (3): 303–311. doi:10.1038/ncb1838. PMC 2748410. PMID 19198600.
- ^ Greco, V.; Hannus, M.; Eaton, S. (2001). "Argosomes: A potential vehicle for the spread of morphogens through epithelia". Cell. 106 (5): 633–645. doi:10.1016/S0092-8674(01)00484-6. PMID 11551510.
- ^ Scott, G.; Leopardi, S.; Printup, S.; Madden, B. (2002). "Filopodia are conduits for melanosome transfer to keratinocytes". Journal of Cell Science. 115 (Pt 7): 1441–1451. PMID 11896192.
- ^ Tilney LG, Portnoy DA (1989). "Actin filaments and the growth, movement, and spread of the intracellular bacterial parasite, Listeria monocytogenes". J Cell Biol. 109 (4 Pt 1): 1597–608. PMC 2115783. PMID 2507553.
{{cite journal}}: CS1 maint: PMC format (link) - ^ a b Robbins JR, Barth AI, Marquis H, de Hostos EL, Nelson WJ, Theriot JA (1999). "Listeria monocytogenes exploits normal host cell processes to spread from cell to cell". J Cell Biol. 146 (6): 1333–50. PMC 1785326. PMID 10491395.
{{cite journal}}: CS1 maint: PMC format (link) CS1 maint: multiple names: authors list (link) - ^ Pust S, Morrison H, Wehland J, Sechi AS, Herrlich P (2005). "Listeria monocytogenes exploits ERM protein functions to efficiently spread from cell to cell". EMBO J. 24 (6): 1287–300. doi:10.1038/sj.emboj.7600595. PMC 556399. PMID 15729356.
{{cite journal}}: CS1 maint: PMC format (link) CS1 maint: multiple names: authors list (link) - ^ Alberti-Segui C, Goeden KR, Higgins DE (2007). "Differential function of Listeria monocytogenes listeriolysin O and phospholipases C in vacuolar dissolution following cell-to-cell spread". Cell Microbiol. 9 (1): 179–95. doi:10.1111/j.1462-5822.2006.00780.x. PMID 17222191.
{{cite journal}}: CS1 maint: multiple names: authors list (link)